Strongylocentrotus purpuratus (Stimpson, 1857)Common name(s): Purple urchin |
|
| Synonyms: |  |
| Phylum Echinodermata
Class Echinoidea Subclass Euechinoidea Order Echinoida Family Strongylocentrotidae |
|
| Strongylocentrotus purpuratus, Little Corona del Mar, CA | |
| (Photo by: Dave Cowles, March 2005) | |
How to Distinguish from Similar Species: Small S. franciscanus can sometimes be purplish, but usually with a tinge of red, and grows much larger than this species.
Geographical Range: Cook Inlet, Alaska to Isla Cedros, Baja California. More abundant in the southern part of its range.
Depth Range: Lower intertidal to 160 m depth
Habitat:_Lower intertidal and subtidal, on pilings. Usually in areas of at least moderate wave action (rarely found in Puget Sound or the inner Straits of Juan de Fuca).
Biology/Natural History: Like all our local urchins, this species eats algae. The teeth grow fast enough in the laboratory to be completely replaced in about 75 days. The species frequently holds rocks, shells, algae, or other debris over itself by its tube feet (picture), or is found in holes eroded into the bedrock, which they apparently dig with their spines and teeth (pictures). In areas where the giant brown kelp Macrocystis grows, that species is the preferred food. They sometimes eat chitons such as Katharina tunicata. In great numbers the urchin can have serious effects on the survival and regrowth of kelp beds after a storm destroys the full-grown kelp. Predators include otters, the seastar Pycnopodia helianthoides (which may swallow them whole), and occasionally the seastars Pisaster ochraceous and Dermasterias imbricata; some crabs, the anemone Anthopleura xanthogrammica, and sheephead fish. The urchin defends itself against seastars by lowering its spines, retracting its tube feet, raising its pedicellariae, and moving to another spot. Sea otter bones are stained purple from the pigment in this urchin's spines. The species tolerates a wide range of temperatures and salinities, from 5 to 23.5C and 80-110% seawater in the lab. Mass mortality occurs if temperatures exceed 26 C. The species requires well-oxygenated water, and obtains oxygen mainly through the tube feet, which are usually extended at least partway when under water. Symbionts include ciliated protozoans and the flatworm Syndisyrinx franciscanus in the gut, and externally the purple polychaete Flabelligera commensalis and the isopod Colidotea rostrata which live among the spines. Most spawning occurs in Jan-March (October-November off northern Baja California), though some reproductive individuals can be found in other seasons. Sexes are usually separate, though some hermaphrodites are found. They are easily induced to shed gametes, which are extensively used to study fertilization and early embryonic development. Urchin eggs fertilized with sand dollar sperm begin development but die as embryos. Off San Diego the gonadal index indicates reproductive activity in spring to summer, with much decreased gonadal index in late fall and early winter. The species grows slowly, with large size reached in about 10 years.
Recent scientific developments (See Scientific Articles section): This species' DNA sequence was reported in 2006 (Sea urchin gene sequencing consortium, 2006). It appears to have about 23,500 genes, similar to the number humans have. Although the larva (pluteus) of this species, which forms by about 2 days after fertilization, has only about 1500 cells of about 12 types, it takes a great deal of gene transcription to get it to that stage. About 11,500 protein-encoding genes and another 51,000 RNA's with some other function had been transcribed by that time. During this time period about 80% of the species' 283 transcription factor genes had already been used at least once. Others were used during development of the egg inside the mother. Together, the use of such a large proportion of the transcription factors so early in development implies that such regulatory genes may be used repeatedly and for different purposes at different stages of life. Other genes heavily involved during development include genes coding for general cytoskeletal and metabolic proteins, while fewer of the genes coding for immunity or sensory functions were transcribed during this period (Samanta et al. 2006).
Even though the urchin has no known organs specialized for light or chemoreception and does not even have a centralized brain, the species has 979 genes associated with sensing light or odors. This includes 6 genes for opsins, which are receptors for sensing light. In the adult at least some of these opsins seem to be concentrated in the pedicellariae and on the tips of the tube feet, while the pluteus has light-sensitive spots on the arms.
The urchin has a remarkably well-developed immune system (Rast et al. 2006). In its innate immune system, which is shared by both vertebrates and invertebrates, it has 218, or more than 10 times as many SRCR genes as do vertebrates and has 222 toll-like receptors compared with 10 for humans. It also has genes which have been associated exclusively with the vertebrate adaptive immune system, such as Rag genes which are involved in producing antibodies in vertebrates (the urchin does not produce antibodies). It also has genes associated with interleukins and tumor necrosis factors, which normally act as signals for vertebrate immune cells which the urchin lacks.
A MRI scan of the internal anatomy of this species, performed by Ziegler et al., (2014), is publically available.
Paganos et al., (2025) reported based on single-nucleus RNA sequencing in the urchin Paracentrotus lividus that, although they have not been considered to even have a central nervous system, nearly the entire nervous system in their body has a genetic activity signature similar to that found specifically in the brain of vertebrates. Only in the gut and vascular system were neurons with activities similar to those in the trunk of vertebrates found. They also discovered that some neurons secrete several different kinds of neurotransmitters, and found 15 different types of photoreceptors. They speculate that this unusual pattern may be true for all urchins. A summary of this article was also published in American Scientist 114:1 (Jan-Feb 2026) p 13.
| Return to: | |||
| Main Page | Alphabetic Index | Systematic Index | Glossary |
References:
Dichotomous Keys:Allen, 1976
Kozloff 1987, 1996
Smith and Carlton, 1975
General References:
Gotshall,
1994
Hinton,
1987
Kozloff,
1993
Lambert
and Austin, 2007
McConnaughey
and McConnaughey, 1986
Morris
et al., 1980
Niesen,
1994
Niesen,
1997
O'Clair
and O'Clair, 1998
Ricketts
et al., 1985
Scientific Articles:
Adams, N.L., J.P. Campanale, and K.R. Foltz, 2012. Proteomic responses of sea urchin embryos to stressful ultraviolet radiation. Integrative and Comparative Biology 52:5 pp 665-680
Basch, L.V. and M.J. Tegner, 1998. Reproductive ecology of sea urchins, Strongylocentrotus purpuratus, at contrasting intertidal and subtidal environments. p. 561 in Rich Mooi and Malcolm Telford (eds), Echinoderms: San Francisco. Proceedings of the Ninth International Echinoderm Conference, San Francisco, California USA 5-9 August 1996.
Bottjer, David J., Eric H. Davdson, Kevin J. Peterson, and R. Andrew Cameron, 2006. Paleogenomics of echinoderms. Science 314: 956-960
Davidson, Eric H., 2006. The sea urchin genome: Where will it lead us? Science 314: 939-940
Ebert, T.A., J.C. Hernandez, and M.P. Russell, 2011. Problems of the gonad index and what can be done: analysis of the purple sea urchin Strongylocentrotus purpuratus. Marine Biology 158:1 pp. 47-58. DOI:10.1007/s00227-010-1541-2
Edmands, S., P. Moberg, and R.S. Burton, 196. Allozyme and mitochondrial DNA evidence of population subdivision in the purple sea urchin, Strongylocentrotus purpuratus. Marine Biology 126: 443-450
Elbert, T.A., S.C. Schroeter, J.D.Dixon, and P. Kalvass, 1994. Settlement patterns of red and purple sea urchins (Strongylocentrotus franciscanus and S. purpuratus) in California, USA. Marine Ecology Progress Series 111: 41-52
Flowers, Jonathan M., Stephen C. Schroeter, and Ronald S. Burton, 2002. The recruitment sweepstakes has many winners: Genetic evidence from the sea urchin Strongylocentrotus purpuratus. Evolution 56:7 pp 1445-1453
Goldstone, JV, A. Hamdoun, B.J. Cole, M. Howard-Ashby, D.W. Nebert, M. Scally, M. Dean, D. Epel, M.E. Hahn, and J.J. Stegeman, 2006. The chemical defensome: Environmental sensing and response genes in the Strongylocentrotus purpuratus genome. Dev Biol. 300(1):366-384
Howard-Ashby M, S.C. Materna, C.T. Brown, Q. Tu, P. Oliveri, R.A. Cameron, and E.H. Davidson, 2006. High regulatory gene use in sea urchin embryogenesis: Implications for bilaterian development and evolution. Dev Biol. 300(1):27-34
Knott, K. Emily, and Gregory A. Wray, 2000. Controversy
and consensus
in Asteroid systematics: new insights to Ordinal and Familial
relationships. American Zoologist 40:3 pp. 382-392
McEdward, Larry R. and Benjamin G. Miner, 2006. Estimation and interpretation of egg provisioning in marine invertebrates. Integrative and Comparative Biology 46:3 pp 224-232
Miller, B.A. and R.B. Emlet, 1997. Influence of nearshore hydrodynamics on larval abundance and settlement of Strongylocentrotus franciscanus and S. purpuratus in the Oregon upwelling zone. Marine Ecology Progress Series 148: 83-94
Paganos, Periklis, Jack Ullrich-Luter, Alba Almazan, Danila Voronov, Jil Carl, Anne-C Zakrzewski, Berit Zemann, Maria Lorenza Rusciano, Tiphane Sancerni, Maria Schauer, Oguz Akar, Filomena Caccavale, Maria Cocurullo, Giovanna Benbenuto, Jenifer Carol Croce, Carsten Luter, and Maria Ina Arnone, 2025. Single-nucleus profiling highlights the all-brain echinoderm nervous system.Sci. Adv.11,eadx7753..DOI:10.1126/sciadv.adx7753Rast, Jonathan P., L. Courtney Smith, Mariano Loza-Coll, Taku Hibino, and Gary W. Litman, 2006. Genomic insights into the immune system of the sea urchin. Science 314: 952-956
Samanta, Manoj P., Waraporn Tongprasit, Sorin Istrail, R. Andrew Cameron, Qiang Tu, Eric H. Davidson, and Viktor Stolc, 2006. The transcriptome of the sea urchin embryo. Science 314: 960-962
Schroeter, S.C., J.D. Dixon, T.A. Ebert, and J.V. Rankin, 1996. Effects of kelp forests Macrocystis pyrifera on the larval distribution and settlement of red and purple sea urchins Strongylocentrotus franciscanus and S. purpuratus. Marine Ecology Progress Series 133: 125-134
Sea urchin gene sequencing consortium, 2006. The genome of the sea urchin Strongylocentrotus purpuratus. Science 314: 941-952
Sodergren, E., Y. Shen, X. Song, L. Zhang, R.A. Gibbs, and G.M. Weinstock, 2006. Shedding genomic light on Aristotle's lantern. Dev Biol. 300(1):2-8
Telford, Malcolm and Olaf Ellers, 1998. The moving teeth of echinoids. pp. 843-848 in Rich Mooi and Malcolm Telford (eds), Echinoderms: San Francisco. Proceedings of the Ninth International Echinoderm Conference, San Francisco, California USA 5-9 August 1996.
Wilt, Fred, Lindsay Croker, Christopher E. Killian, and Kent McDonald, 2008. Role of LSM34/SpSM50 proteins in endoskeletal spicule formation in sea urchin embryos. Invertebrate Biology 127(4): 452-459
Wray, Gregory A., 2006. The evolution of embryonic gene expression in sea urchins.
Ziegler, Alexander, Cornelius Faber, Susanne Mueller, Nina Nagelmann, and Leif Schroder, 2014. A dataset comprising 141 magnetic resonance imaging scans of 98 extant sea urchin species. GigaScience 2014, 3:21 doi: 10.1186/2047-217X-3-21 (Open access)
General Notes and Observations: Locations, abundances, unusual behaviors:

Strongylocentrotus purpuratus (left) is
much
smaller
than its
relative Strongylocentrotus
franciscanus (right), and has shorter
spines. Photo by Dave
Cowles, July 2005

The spines of S. purpuratus are only about 2 cm or
less long,
as contrasted with the longer spines of S.
franciscanus.
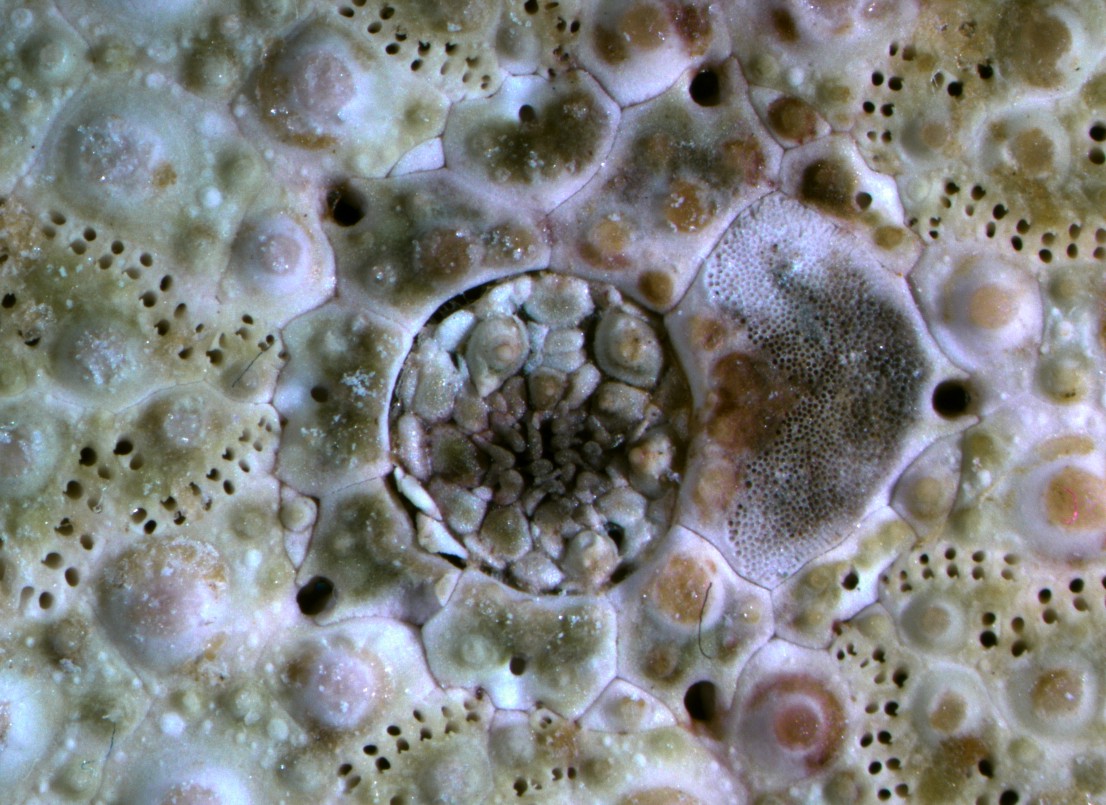
This view shows the intricate network of plates and openings on the aboral side of the urchin. The periproct plates in particular are weakly held together and often collapse after the animal dies. The 5 gonopores are located in 5 genital plates arranged around the periproct. One of the genital plates is also the madreporite. Clicking the image will take you to a larger, unlabeled version. Photo by Dave Cowles, Oct 2012.

This species is often found within holes they grind
in
rock
(probably
over multiple generations) with their teeth and spines. They
may
even erode holes in steel pilings by flaking off rust.
The photos above and below were taken at Little Corona del Mar, CA
by Dave Cowles, March 2005


The species often covers itself with rocks and algae, which it holds
onto with its tube feet, as seen in this tidepool at Dana Point, CA
Photo by Dave Cowles, March 2005

These individuals in a Cape Flattery tidepool have covered themselves with fragments of shells and dead barnacles. Photo by Dave Cowles, July 2010

This deformed test of a purple urchin was found washed up on the beach
at Slip Point, WA in July, 2018. Photo by Dave Cowles
Authors and Editors of Page:
Dave Cowles (2005): Created original page