Hyperoche medusarum (Kroyer, 1838)Common name(s): |
|
| Synonyms: Metoecus abyssorum, Metoecus medusarum, Tauria kroeyeri, Tauria tauriformis, Tauria prehensilis, Tauria abyssorum, Tauria luetkeni |  |
|
Phylum Arthropoda
Subphylum Crustacea
Class Malacostraca
Subclass Eumalacostraca
Superorder Peracarida
Order Amphipoda
Suborder Hyperiidea
Infraorder Physocephalata
Superfamily Phronimoidea
Family Hyperiidae
|
|
| A male Hyperoche medusarum riding on the ctenophore Pleurobrachia bachei. The amphipod is 5 mm long and is facing away from the ctenophore, hanging onto it like Atlas holds the world. Note the long antennae. | |
| (Photo by: Dave Cowles, August 2014). My thanks to Joanna Cowles for discovering this amphipod. | |
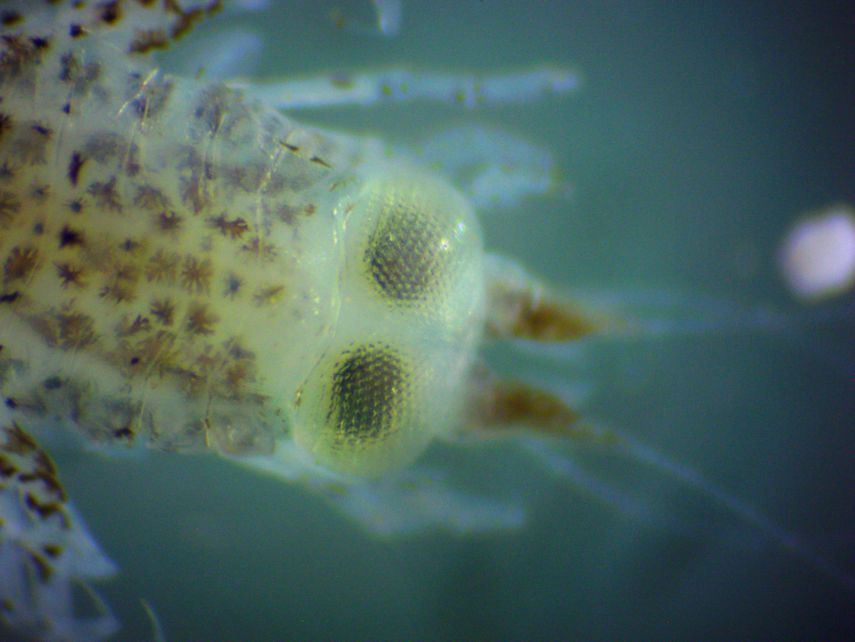
Description: Members of Infraorder Physicephalata usually have a pereon which is not spherically inflated and is about the same width as the pleon. They usually have large eyes. The antennae of males and females differ in shape. Pereopods 1 and 2 are usually chelate or subchelate. Members of Superfamily Phronimoidea have antennae which extend from the front of the head. Antennae are short in females but long in males. Members of Family Hyperiidae usually have dark pigment, and a head which is large and rather spherical, larger than the first somite of the pereon. The eyes occupy much of the head and touch each other dorsally (photo). The uropods have free rami (photo). Members of genus Hyperoche have all pereonsomites and coxal plates free. The head is large and semicircular. The eyes cover the entire dorsal and lateral surfaces of the head. Antenna 1 has 3 segments in its peduncle, and in females it is longer than antenna 2. Antennae 2 have 4 segments in the peduncle in females. The chelae on pereopods 1 and 2 are formed from segments 5 & 6 of the legs, rather than segments 6 & 7. The 7th segment is small and projects from the chela. The fifth segment of pereopods 1 & 2 (the fixed claw of the chelae) is laterally compressed, knife-shaped, and has small teeth along the anterior margin. Hyperoche medusorum is the type species for this genus. It has the posterior distal angle of epimerons 2 & 3 slightly acute rather than rounded or straight. The anterior distal angle of the 6th segments of pereopods 1 & 2 is not stretched out above the claw. The posterior margin of the 5th & 6th segments of pereopods 3 & 4 have small teeth. Total length usually not more than 5-6 mm but up to 15 mm for females.
How to Distinguish from Similar Species: H. luetkenides has setae but not teeth on the posterior margin of the 5th & 6th segments of pereopods 3 & 4.
Geographical Range: Bipolar: In the North Pacific from Japan through the Okhotsk and Bering Seas and Alaska down to the Gulf of California. From the Arctic Ocean it extends into the North Atlantic down to Ireland and Labrador and to Chesapeake Bay. In the Southern Ocean it has been found near the South Georgia Islands, SW Africa south of 25 degrees S, off Argentina, and off southern New Zealand.
Depth Range: Epipelagic (upper 30 m), but caught at 1100 m in the Gulf of California (Gasca and Haddock, 2004)
Habitat: Parasitic on gelatinous organisms
Biology/Natural History: Adults of this species parasitizes several species of ctenophores such as Pleurobrachia and Mnemiopsis, as well as medusae such as Cyanea capillata. One of the prey items consumed by the free-swimming juveniles are fish larvae such as Pacific Herring (Westerhagen and Rosenthal, 1976). The juvenile amphipods can occur in high numbers so their impact on the fish larvae may be substantial. Juveniles can often be found sitting on the exumbrella of hydromedusae such as Tiaropsis, Sarsia, Phialidium, and Polyorchis.
Some authors claim that hyperiid amphipods are second in numbers in the plankton only to copepods and euphausiids.
Merresunters (1976) gathered large numbers of juveniles at the Pacific Biological Station, Nanaimo, Vancouver Island, Canada by suspending a 300 watt bulb over the water at the station dock at night and scooping them up as they aggregated in the light. The species is common in Departure Bay (near Nanaimo) in April and May.
Spicer and Morritt (1995) studied the blood of this and several other species of Hyperiidean amphipods and concluded that their blood contains no hemocyanin.
Adults of Family Hyperiidae usually are strongly sexually
dimorphic
| Return to: | |||
| Main Page | Alphabetic Index | Systematic Index | Glossary |
References:
Dichotomous Keys:Vinogradov et al., 1982
General References:
Scientific Articles:
Bowman, T.E., 1953. The systematics and distribution of pelagic amphipods of the families Vibiliidae, paraphronimidae, Hyperiidae, Dairellidae, and Phrosinnidae from the northeastern Pacific. Ph.D. thesis, University of California, Los Angeles. 430 pp.
Bowman, T.E., C.D. Meyers, and S.D. Hicks, 1963. Notes on associations between hyperiid amphipods and medusae in Chesapeake and Narranansett Bays and the Niantic River. Chesapeake Science 4: pp 141-146
Gasca, Rebeca and Steven H.D. Haddock, 2004. Associations between gelatinous zooplankton and hyperiid amphipods (Crustacea: Peracarida) in the Gulf of California. Hydrobiologia 530/531: pp 529-535
Hurley, D.E., 1956. Bathypelagic and other Hyperiidae from Californian waters. Allan Hancock Foundation Occasional Paper 18. 25 pp.
Laval, P., 1980. Hyperiid amphipods as crustacean parasitoids associated with gelatinous zooplankton. Oceanography and Marine Biology Annual Review 18: pp 11-56
Lorz, H.V. and W.G. Pearcy, 1975. Distribution of hyperiid amphipods off the Oregon coast. Journal of the Fisheries Research Board of Canada 32: pp 1442-1447
Sorarrain, Dora R., Fernando Ramirez, and Hermes Mianzan, 2001. Hyperoche medusarum (Kroyer, 1838) (Amphipods, Hyperiidae) and Mnemiopsis McCradyi (Mayer, 1910) (Ctenophora): a new host and first record of this association for the southwestern Atlantic. Crustaceana 74:4 pp 407-410
Spicer, John I. and David Morritt, 1995. Oxygen carriage by the haemolymph of Hyperiid Amphipods. Journal of the Marine Biological Association of the United Kingdom 75:4 pp 997-998
Westerhagen, H. von, 1976. Some aspects of the biology of the hyperiid amphipod Hyperoche medusarum. Helgolander wissenschaft Merresunters 28: pp 43-50
Westerhagen, Hein von, and Harold Rosenthal, 1976. Predator-prey relationship between Pacific Herring, Clupea harengus pallasi, larvae and a predatory Hyperiid amphipod, Hyperoche medusarum. Fishery Bulletin 74:3 pp 669-674
Westerhagen, H. von, H. Rosenthal, and G. Furstenberg, 1979. Factors influencing predation of Hyperoche medusarum (Hyperiida: Amphipoda) on larvae of the Pacific Herring Clupea harengus pallasi. Marine Biology 51: pp 195-201
Web sites:
General Notes and Observations: Locations, abundances, unusual behaviors:

This closeup photo shows how the amphipod is hanging onto the
ctenophore,
between two ctene rows.

Dorsal view after the amphipod was removed from its ctenophore host

| Dorsal and ventral views of the head | |
 |
 |
| In dorsal view, the fact that the eyes occupy the entire dorsal head can be seen. | In this ventral view they large eyes which wrap around the entire lateral sides of the head can be seen. Also visible is the mouth, plus the chelae on the first and second pereopods, which are composed of leg segments 5 & 6 rather than 6 & 7. The 7th leg segment (dactyl) can be seen as a narrow extension at the end of the chela. |

A view of the uropods
Authors and Editors of
Page:
Dave Cowles (2014): Created original page
CSS coding for page developed by Jonathan Cowles (2007)
Salish Sea Invertebrates web site provided courtesy of Walla
Walla University